
Gymnosperms
Definition
of Gymnosperms:
The
term gymnosperms (gymnos = naked; sperma = seed) was introduced by Theophrastus
in 300 BC
to
describe plants with unprotected seeds. Gymnosperms are phanerogams without
ovary.
The
phanerogams or Spermatophyta (sperm = seed; phyton = plant) or seed plants
are those plants which reproduce by means of seeds, not spores. Gymnosperms are
the vascular plants where seeds are not enclosed within an ovary (opposite to
an angiosperm or flowering plants where seeds are enclosed by mature ovaries or
fruits). In these plants the ovules are borne naked or the surface of the
megasporophylls,
which are often arranged in the cones. Fossil records indicate that the
gymnosperms must have evolved approximately 300 million years ago
from
non-seed producing ancestors of the extinct division of Progymnospermophyta
which were fern like in appearance (form a bridge between pteridophytes and
angiosperms). Gymnosperms were dominant plants over the earth’s surface during
the jurassic and cretaceous periods of mesozoic era. At present about 83 genera
and approximately 790 species of living gymnosperms are
distributed
throughout temperate, tropical and arctic regions of the world.
Characteristics of Gymnosperm:
Gymnosperms
are, those seed plants in which the seeds remain exposed over the surface of
the megasporophylls because the latter are not folded to form pistils.
1.
Gymnosperms are a small group of seed plants which are represented by only 900
living species.
2.
Gymnosperms are more ancient than the angiosperms. They formed dominant
vegetation on earth some 200 million years back in mesozoic era. Today they are
dominant only in cold areas, where instead of rain; snow is the source of
water.
At
other places they have been replaced by angiosperms. In warmer areas only a
handful of gymnosperms can be observed, e.g., Cycas (like C. circinalis in
South India), Araucaria (native of South America, New Zealand and East
Australia, like A. heterophylla.
3.
All gymnosperms are perennial and woody, forming either bushes or trees. Some
of these are very large and live for thousands of years, e.g., Sequoia sempervirens (tallest gymnosperm
of 111.6 m) Zamia pygmia is smallest
(26 cm).
4.
Flowers are absent. Two types of sporophylls, microsporophyll’s and megasporophylls
are usually aggregated to form distinct cones or strobili, pollen cones (male
cones) and seed cones (female cones) respectively.
5.
Seeds do not occur inside a fruit. They are naked.
6.
A distinction of ovary, style and stigma is absent.
7.
Ovules are orthotropous and sessile. Each ovule is surrounded by a 3-layered
integument.
8.
Female gametophyte contains archegonia.
9.
Pollination is direct as a stigma is absent and the pollen grains directly
reach the micropylar ends of Ovules. Pollination is usually accomplished by
wind (anemophily).
10.
Male gametophyte produces only two male gametes or sperms. Generally one of
them is functional.
11.
An external water is not required for
transport of male gametes. Instead, a pollen tube is formed by the male
gametophyte for effecting fertilization (siphonogamy).
12.
Seeds contain a food laden tissue or endosperm for future growth of embryo into
seedling. The tissue represents the female gametophyte.
13.
Like pteridophytes, xylem does not possess vessels except in some gneophytes.
Phloem is without
companion
cells and sieve tubes. Sieve cells are not arranged end to end in rows.
14.
Vascular tissues are arranged into vascular bundles just like angiosperms.
Vascular bundles of stem are open so that secondary growth is quite common.
Classification of Gymnosperm as per
Sporne (1965):-
This
is a recent classification. The gymnosperm are considered as division.
According to the Sporne the gymnosperms are divided in to three classes which
include both fossil and living genera. The three classes are, cycadopsida,
conoferopsida and gnetopsida. Each class is further divided into orders and
each order again divided into families. The outline of Sporne’s classification
is given. below,

1.
Class: Cycadopsida:- The plants are tree sized, stem unbranched, leaves are
pinnately compound i.e palm like. The wood is porous and loosely arranged i.e
monoxylic. Seed is radially symmetrical. This class is further divided in to
four orders namely Pteridospermales, Bennetitales, Pentoxylales are represents
fossil genera and cycadales have both fossil & living genera. e.g Nilsonia
(fossil genus), Cycas, Zamia, Ceratozamia. Each order further divided into families.
2.
Class: Coniferopsida:- Plants are large tree, stem is branched, leaves are
simple. The wood is compact called pycnoxylic. This is further divided into
four orders namely cordaitale is fossil order and remaining coniferales,
taxales and ginkgoales are living as well as fossil.
The
order coniferale is important represents nine families out of that family
pinaceae represent genus Pinus. The Trichopytis is a fossil genus and Taxus is
a living genus. The Ginkgo is living fossil
3.
Gnetopsida:- The plants are median tree, shrub or woody climbers. Leaves are
simple, with opposite or whorled phyllotaxy. The xylem vessels are present is
important character of this class. They have only one gnetales order and three
families; out of that Gnetaceae represents Gnetum. The Gnetum occurs in western
ghat of Maharashtra, The genus Ephedra in Rajasthan & U.P. The genus
Welwitschia is not found in India.
Economic Importance of Gymnosperm:-
1.
The Pinus wood is soft therefore used for boxes, furniture, poles and toys.
2.
The Juniperous wood is used for making boat, furniture and making lead pencil.
3.
Abies and Picea used for making cabinets and small boxes.
4.
The resin obtained from Pinus and Agathis used in manufacturing paints &
varnishes.
5.
Turpentine of Pinus is used in inks, adhesives and some medicines.
6.
Canada balsam obtained from Abies balsamea is useful mounting medium for
biological preparations.
7.
The gum is also obtained from Cycas and Araucaria.
8.
The aromatic oil obtained from many species used in making perfumery product.
9.
The seeds of Pinus & Gnetum are edible and tuber of Zamia are edible.
10.
The starch obtained from Cycas is used for manufacturing Sago.
11.The
young, delicate leaves of Cycas & Gnetum are used as vegetable.
12.Cedarwood
oil obtained from Cedar have god medicinal value.
13.The
drug ephedrine obtained from Ephedra is useful for treatment of asthmas and
cold.
14.The
stem extract of Cycas pectinata is used as hair wash. The leaf juice of Cycas
circinalis is used for stomach disorders, vomiting and skin diseases.
15.Cycas
pollen grain have nacrotic property which is medicinally useful.
16.The
tannin obtained from Tsuga and Picea species is used in tanning industry.
17.Thuja,
Cycas, Araucaria (Christmas tree) cultivate in garden and Parks as ornamental.
18.Fibres
obtained from the Cycas revolata and Gnetum bark used in making rope & mat.
19.The
crushed seeds of C.circinalis are used as fish poison.
20.The
coniferous plant check the soil erosion and bind the soil. Therefore used for
plantation
on hill slopes.
Cycas
Distribution of Cycas:
It is the only genus of the family Cycadaceae, which is represented in
India. It has got about 15 species (Sporne, 1965), 20 species (Willis, 1966)
which are widely distributed in Eastern as well as Western hemisphere from
Madagascar, Eastern coast of Africa to Japan and Australia touching China and
India.
In our country a few of the species are found growing abundantly in the
South Andaman and Nicobar islands, Madras, Mysore, Malabar and in North East in
Bengal, Assam, Nepal and Sikkim. A few of the species are also found in Burma
and Ceylon.
.
The following species are found in
India:
C. circinalis:
Plants are about 12 to 15 feet tall. Leaves are 5 to 9 feet long. It is
distributed in western part of peninsular India, Malabar. Orissa hills, Andhra,
Madras to Ceylon upto 3500 ft. The plants are also cultivated in Indian
gardens. In Hindi it is called as Jangli-Madan mast-ka-phul.
C. pectinata:
.
Plants are about 8 to 10 feet tall. Leaves are 4 to 6 feet long. It is
commonly found growing in Nepal. Sikkim, in Assam Khasia Hills, East Bengal and
Burma. In Nepal it is commonly called as Thankal.
C. beddomei:
Plants are about 40 cm. tall. Leaves 3 feet long. It is naturally found
growing on dry hills of Cuddapah in Andhra Pradesh, Madras, Malabar Java.
Locally it is called as Per-ita.
C. rumphii:
Plants are more than 12 feet tall. Leaves 4 to 5 feet long. It is found
growing in Andaman and Nicobar islands, Burma etc. It is also cultivated in
Indian gardens. In Tamil it is called as Kama, Paiyindu.
C. revoluta:
Plants are upto 10 feet tall. Leaves 3 to 5 feet long. It is a native of
China and Japan and locally called as Tesso. In our country it is mostly
cultivated in gardens and locally called as Sago palm. Due to its primitive
characters it is also known as living fossil. Exceptionally it has reached a
height of about 20 feet in gardens of Taj Mahal Agra (India).
.
C. siamensis:
Trunk geophilous, tuberous but at times 6 feet tall.Leaves upto 4 feet
long. It is a native of Burma, Cochin, China etc. Locally in Burma it is called
as Mondaing. Pant and Nautial (1963) considered C. siamensis end C. pectinata
similar on the basis of epidermal and anatomical studies.
Morphological Features of Cycas:
Cycas is perennial, slow growing evergreen plant and is referred as
living fossil because it occurs as a fossil e.g., C. fusiana. It looks like a
palm tree. Its main plant body is sporophytic, diploid, dominant and can be
differentiated into three parts – roots, stem and leaves. Tallest species of
Cycas is C. media with 20 feet height.
1. Roots:
They are of two types – normal and coralloid roots. Normal roots grow
deep into the soil and form tap root system. Later it is replaced by
adventitious roots. The function of these roots is to fix the plant in the soil
and to absorb water and other minerals.

From the normal roots develop some small lateral apogeotropic branches
near the ground surface. These lateral roots get infected with bacteria, fungi
as well as algae. The entry of these organisms is said to be responsible for
the characteristic, swollen, knob like or coral like appearance and hence,
these roots are called as coralloid roots or corallorhiza. These roots have
minute pores (lenticels like) which are respiratory in function (aeration).
Root cap and root hairs are absent in coralloid roots (Fig. 2).

2. Stem:
It is thick, erect, woody, aerial and usually unbranched (caudex).
Branching is rare and it is due to injury or development of adventitious buds.
Surface of the stem is rough due to the presence of persistent woody leaf bases
(Fig. 4). These leaf bases form thick armour around the stem.
In the armour are distinctly visible the alternating bands of large and
small rhomboidal leaf bases. Larger ones are of foliage leaves and smaller ones
are of scaly leaves and megasporophylls in the female plant. The leaf bases are
spirally and compactly arranged with each other (Fig. 4).

At the top is present a crown of leaves (Fig. 1).
3. Leaves:
Leaves are dimorphic i.e., of two types – scale leaves and foliage
leaves. Both these types of leaves form a crown at the top of the stem.
(a) Scale leaves:
These are small, dry, brown, triangular structures with a thick covering
of brown hairs or rameta. These leaves alternate with green foliage leaves.
These leaves protect the shoot apex and reproductive structures (Fig. 3).

(b) Foliage leaves:
These leaves are also produced in a crown at the apex of the stem.
According to Coulter and Chamberlain (1910) one crown of foliage leaves is
formed yearly while D.D. Pant (1953) observed the formation of two crowns per
year in C. circinalis. In case of C. revoluta the leaves are 30 to 150 cm long
but in case of C. circinalis they are up to 270 cm long.
A single foliage leaf is pinnately compound. It is unipinnate and
paripinnate. Each leaf has 80-100 pairs of leaflets which are arranged on both
the sides of adaxial groove of the rachis in opposite or alternate manner. The
rachis is spiny below with the sheathing leaf base (Fig. 6A). these spines are
modified leaflets. Each leaflet is leathery in texture, sessile elongated,
ovate or lanceolate in shape and has entire margin with acute apex. Each pinna
or leaflet contains a midrib without lateral veins.
In C. micholitzii the leaflet is repeatedly and deeply dichotomised
(Fig. 5).

Margins of the pinnae are flat (Fig. 6B) but sometimes they are curved
downwards and inwards (revolute) (Fig. 6C) which give the plant a specific name
C. revoluta. Young leaves have circinately coiled leaflets which are also
covered by hairs or ramenta like those of ferns (Fig. 6 D, E).

Internal Structure of Cycas:
1. Root:
(i) Normal root:
Its internal structure is exactly similar to that of dicot root. It is
circular in outline and can be differentiated into epiblema, cortex and
vascular tissue.
a. Epiblema:
It is the outermost limiting layer and consists of single layer of thin
walled cells. Some of its cells give rise to root hairs.
b. Cortex:
Epiblema surrounds the multilayered zone of thin walled parenchymatous
cortex with numerous intercellular spaces. The cells of the cortex are filled
with starch. Some tannin cells, mucilage cells and sometimes sphaeraphides
(calcium oxalate crystals) are also present in the cortex. The innermost layer
of the cortex forms the endodermis which is characterised by the presence of
casparian strips.
c. Vascular tissue:
Endordermis is followed by multilayered parenchymatous pericycle.
Vascular bundles are radial. Xylem is diarch and exarch i. e., protoxylem is
towards the periphery). The protoxylem consists of spiral tracheids whereas the
metaxylem consists of scalariform thickenings. Vessels are absent. Alternating
with the protoxylem groups are present phloem cells consisting of sieve tubes
and phloem parenchyma. The companion cells are completely absent (Fig. 7A, B).

Secondary Growth:
The mature normal root shows secondary growth on both the lateral sides
of primary xylem. Along with the inner side of primary phloem develops the
cambium. It cuts off secondary phloem on outer side and secondary xylem on the
inner side. After sometime the cells of the pericycle opposite to the
protoxylem strands also become meristematic and behave as cambium, cutting
phloem on the outer side and xylem on the inner side.
Thus, a complete ring of cambium is formed which forms a complete ring
of secondary xylem on the inner side and complete ring of secondary phloem on
the outer side. The primary phloem is crushed in the due course of development
and appears in the form of crushed layer above the secondary phloem.
Simultaneously the formation of periderm also starts. The cells of the
outermost layer of the cortex become meristematic (also called cork cambium)
and start cutting cork cells on the outer side and secondary cortex on the
inner side. In the course of the formation of cork, the cells of the epiblema are
crushed (Fig. 8 A, B)

(ii) Coralloid Root:
The transverse section of the coralloid root is similar to that of
normal root and it can be differentiated into epidermis, cortex and vascular
tissue.
a. Epidermis:
In young root, it is similar to normal root. However, in old root the
outermost tissue is periderm. It consists of 2 to 5 layers of dead cells.
b. Cortex:
The cortex is wider in comparison with the normal root. A greenish algal
zone is present almost in the middle of the cortex and divides it into outer
cortex and inner cortex (Fig. 9A, B).
The algal zone consists of loosely connected, radially elongated thin
walled cells occupied by blue green algae (Anabaena cycadae, Nostoc
punctiforme, Oscillatoria), bacteria (Azotobacter, Pseudomonas radicicola) and
some fungi. The main function of these roots is nitrogen fixation due to the
presence of cyanophycean members. Endodermis is similar to normal root.

c. Vascular tissue:
Endodermis is followed by multilayered parenchymatous pericycle.
Vascular bundles are radial. Xylem is triarch and exarch.
Secondary growth is very rare or absent. No secondary xylem or secondary
phloem are developed although cork and cork cambium are present.
Comparison between Normal Root and
Coralloid Root:
Normal root:
1. Develops from the radicle, tap root system
2. Geotropic
3. Such characters are absent
4. Such infection is absent
5. Root hairs are present
6. Cortex is smaller
7. Such division is absent
8. Diarch
9. Secondary growth present
10. Main functions are: fixation of plant, absorption of water and
mineral nutrients
Coralloid root:
1. Develops from the normal roots
2. Apogeotropic
3. Develops from the normal roots
4. Dichotomously branched and appears like coral
5. Gets infected with algae, bacteria and fungi
6. Absent
7. Cortex is wider in comparison
8. Due to presence of the algal zone in the cortex, it is differentiated
into outer cortex and inner cortex
9. Very little or absent
10. Main function is nitrogen
fixation
A transverse section of young stem is similar to dicot stem. It is
irregular in outline due to persistent leaf bases. Internally, it can be
differentiated into epidermis, cortex and vascular cylinder.
a. Epidermis:
It is the outermost layer of the stem. It is made up of compactly
arranged thick walled cells. Epidermis is ruptured due to the armour of
persistent leaf bases (Fig. 11A).

b. Cortex:
Epidermis encloses the cortex. It forms the major portion of the stem.
It is composed of parenchymatous cells which are filled with large number of starch
grains. These starch grains are the source of sago starch. Therefore, C.
revoluta is popularly known as sago palm.
Scattered in the cortex are various mucilage canals. Each mucilage canal
is lined by many radially elongated epithelial or secretory cells (Fig. 10).
which secrete mucilage. These canals are connected with those of the pith with
the help of the medullary rays. The innermost layer of cortex is endodermis. It
is not distinct.

c. Vascular Cylinder:
The vascular cylinder is surrounded by not very conspicuous pericycle.
Like dicot stems vascular cylinder consists of many conjoint, collateral, open,
endarch vascular bundles arranged in a ring (ectophloic slphonostele). The
xylem consists of tracheids and Xylem parenchyma (Fig. 11B).
Vessels are absent. Outside the xylem is the phloem which consists of
sieve tubes and phloem paraenchyma. Companion cells are absent. The Xylem is
separated from the phloem with the help of primary combium. The cells of the primary
cambium are brick shaped.
The cells lying in between the vascular bundles form the medullary rays.
These are parenchymatous and connect the pith with the cortex. Each medullary
ray is one celled wide and 1 to 20 cells long.
3. Pith:
In the centre of the stem is present large canals leaf traces massive
pith consisting of parenchymatous cells which are rich in starch (sago starch).
A large number of mucilage canals are also present, which are exactly similar
in structure with the mucilage canals present in the cortex.
4. Leaf Traces and Girdle Traces:
The leaf traces are scattered in the cortex of the stem and constitute
the vascular tissue of the leaves from the main vascular cylinder. Each leaf
receives four traces, two of which are direct traces and the other two are
given out from the opposite side of the direct traces.
These two traces take a round around the main vascular cylinder in
opposite direction through cortex and then enter the leaf base of opposite side
from the point of their origin from the stele. These leaf traces are known as
girdle traces or indirect trances and are peculiar structures in the stem of
Cycas.

Secondary growth:
It is a slow process. At first a complete ring of cambium is formed by
the development of interfascicular combium in between the adjacent vascular
bundles. The cambium cuts off secondary xylem on the inner side and secondary
phloem on the outer side.
Tracheids consist of multiseriate bordered pits. This cambial ring is
short-lived and new cambial ring is formed every year in the pericycle of the
cortex. Wood formed by this method (more than one) cambium ring is polyxylic
and manoxylic (large amount of parenchyma is cut off in the xylem. (Fig. 13).

5. Rachis:
A transverse section of the rachis is somewhat rhomboidal in outline,
but a little higher up it is shield shaped. Its internal structure can be
differentiated into epidermis, cortex and Vascular bundles.
a. Epidermis:
It is the outermost covering. It is made up of compactly arranged thick
walled cells. It is single layered, covered with thick cuticle and has stomata.
Hypodermis:
Epidermis is followed by hypodermis. It is differentiated into outer
2-3- layers of chlorenchyma (Chlorophyll containing thin walled cells) and
inner 4-6 layers of sclerenchyma (thick walled, lignified cells; Fig. 14A, B).
Ground tissue:
Below the sclerenchyma is present a large tissue made up of thin walled
parenchymatous cells. It is called ground tissue. In this region are present
many mucilaginous canals and vascular bundles.
b. Vascular bundles:
Vascular bundles are arranged in the shape of inverted Greek letter
‘omega’ [Ω; Fig. 14 A], Each vascular bundle is conjoint, collateral, endarch,
open and diploxylic i. e., consists of centripetal and centrifugal Xylem and is
surrounded by bundle sheath.
Xylem is present towards the inner side and consists of tracheids and
xylem parenchyma. Vessels are absent. Phloem is present towards the outer side
of the vascular bundle. It consists of sieve tubes and phloem parenchyma.
Companion cells are absent, Cambium is present in between the xylem and phloem.

In rachis the vascular bundles are endarch at the base (centrifugal
xylem is well developed, protoxylem faces towards the centre showing endarch
condition, centripetal xylem is not developed), mesarch in the middle
(centripetal and centrifugal xylem are present showing diploxylic condition)
and exarch at the apex (centripetal xylem is well developed, triangular and
exarch, centrifugal xylem is much reduced and in the form of two patches lying
one on each side of the protoxylem elements of centripetal xylem) due to twisting
of the rachis (Fig. 15 A-C).

6. Leaflet:
The leaflet of Cycas is dorsiventral and hypostomatic (the stomata are
present at the lower surface only). In a transverse section the leaflet can be
differentiated into a swollen midrib portion and two lateral wings (Fig. 16A,
B).
Its internal structure is as follows:

a. Epidermis:
It is the outer most single layer made up of squarish cells. The upper
epidermis is complete whereas the lower epidermis is interrupted by several
sunken stomata present in the region of the wings. The upper and lower
epidermis is covered by a thick layer of culicle.
b. Hypodermis:
Below the epidermis occurs the thick walled sclerenchymatous hypodermis.
It is single layered in the region of blade but in the region of mid rib it
becomes 2-3 layered thick. Two to five layers of sclerenchymatous cells are
also present above the lower epidermis only in the region of the mid rib. It
helps in checking the rate of transpiration and protects the tissue from
excessive heat.
c. Mesophyll:
A well-developed mesophyll tissue is present in the leaflet. It is
differentiated into palisade tissue and spongy parenchyma. Palisade tissue is
present in the form of continuous layer below the sclerenchymatous hypodermis.
Spongy parenchyma present only in the wings directly above the lower epidermis.
It is made up of loosely arranged oval cells filled with chloroplast. These
cells have many intercellular spaces filled with air.
d. Vascular bundle:
A single large vascular bundle is present in the mid rib region of the
leaflet. It is surrounded by a single layer of sclerenchymatous cells, known as
bundle sheath. The vascular bundle is conjoint, collateral, open and
diploxylic. Xylem is present towards the dorsal surface and phloem is present
towards the ventral surface.
Xylem and phloem are separated by a non-functional strip of cambium.
Centrifugal xylem is represented by two small groups on either side of the
protoxylem. The remaining space of the vascular bundle is filled with thin
walled parenchymatous cells.
e. Transfusion tissue:
Groups of tracheidal cells, separated by some parenchymatous cells, or
directly in contact with the centripetal xylem, the bundle sheath are present
in the leaflet. It is called primary transfusion tissue. The cells of this
tissue are short and wide with are reticulate or bordered pitted walls.
A zone is present on either side of the midrib between the palisade and
spongy layers. It is three layered and is composed of elongated colourless
cells. These cells run paralled to the leaf surface from the midrib to the
margin. This zone is called accessory transfusion tissue or secondary
transfusion tissue or hydrostereom or radial parenchyma.
On either side of the leaflet it is connected with the primary
transfusion tissue present around centripetal xylem of the vascular bundle.
Primary and secondary transfusion tissue help in the lateral conduction of
water. The presence of transfusion tissue is to compensate for the unbranched
condition of the midrib and it probably serves as a later conducting channel of
water.
Reproduction in Cycas:
Cycas reproduction by two method – Vegetative and Sexual
1. Vegetative reproduction:
It is the simplest method of reproduction. It takes place by the
formation of bulbils or adventitious buds. These buds develop on the stem in
the axil of the scale leaves. A bulbil is an oval structure, broad at the base
and pointed at the apex. It consists of dormant stem in the centre covered by
numerous brown scaly leaves.
On detachment from the stem, a bulbil starts to germinate by producing
many roots from the lower side and a leaf towards the upper side. A bulbil from
male plant will develop only into male plant while the bulbil from the female
plant will form only female plant because cyas is strictly dioecious (Fig. 1A,
B).
2. Sexual Reproduction:
Sexual reproduction in cycas is
oogamous (the female gamete i.e., egg cell is significantly larger than the
male gamete and is non-motile). Cycas is sporophytic and strictly dioecious
i.e., male and female sex organs are borne on separate plants.

Male Reproductive Organs:
Male plant of cycas produces every year a single male cone (Fig. 1B) at
its apex. In the formation of the male cone the apical meristem is used up, and
therefore, the growth of the steam checked for some time some time but later an
apical meristem is formed at the base of the cone, which pushes that on one
side so that the growth of the stem is resumed again.
Such growth of the stem is called sympodial (Fig. 18 A, B). The male
cone is largest in the plant kingdom (approximately 500 cm or more in length).

Longitudinal Section of Male Cone:
Each cone is an ovoid or conical structure (Fig. 19A). A longitudinal
section of male cone shows that each one consists of a central axis around
which, a large number of leaf like structures called as microsporophylls are
attached at right angle in a compact, spiral, acropetal succession (Fig. 19.B).
The maturation of the sporophylls takes place in a spiral manner i.e., from
apex to base. However, a few sporophlills at the apex and base remain sterile.
Fig. 19 B.

Structure of microsporophyll:
Each microsporophyll represents a stamen. It is a flattened, woody and
triangular structure. It is differentiated into upper or distal, sterile region
called apophysis (Fig. 20 A) and proximal wedge shaped fertile part. Each
microsporophyll bears several hundred microsporangia (pollen sacs) on its
abaxial surface (more than 1000, Fig. 20B-D).
Microsporangia are arranged in clusters of 3 to 6. Each cluster or group
of microsporangia is called sorus. In between the microsporangia indusial hairs
are present which help in the dispersal of the microspores and protect young
sporangia.

Development of Microsporangium:
The development of microsporangium is of eusporangiate type i. e.,
develops from a group of cells called microsporangial initials (Fig. 21A).
These cells are hypodermal in origin and divide by periclinal walls into upper
primary wall cells and lower primary sporogenous cells.
Primary wall cells divide and redivide to form three to six layered wall
below the epidermis. Simultaneously primary sporogenous cells also divide and
redivide irregularly to form a mass of cells known as sporogenous tissue.
At this time from the peripheral cells of the sporogenous tissue or the
inner most wall layer differentiates into a single celled, nourishing layer
known as tapetum (Fig. 21C, D). The outermost layer of the sporangial wall
forms the epidermis or exothecium.
The rest of the sporogenous tissue increases in size and functions as
spore mother cells. These are the ultimate cells of sporophytic phase. Each
spore mother divides cell by reduction division to form four haploid
microspores (Fig. 21D-F).

A mature microsporangium is sessile or shortly stalked consists 5-6 wall
layers (outer single layer exothecium, innermost single layer tapetum and rest
endothecial layers). The wall of sporangium encloses large number of haploid
microspores. The tapetum is used up during the development of microspores. Only
the signs of disintegrated tapetum can be seen at maturity.
Female Reproductive Organs:
Female reproductive organs are megasporophylls. Each female plant every
year produces numerous megasporophylls in acropetal succession above each crown
of foliage and scaly leaves. There is no female cone formation. The number of
the megasporophylls is much more than the number of the foliage leaves on the
stem.
During the formation of the megasporophylls the apical meristem is not
used up like that of male cone and therefore, the growth of the stem continues,
and thus in female plant growth is monopodial.
Structure of Megasporophyll:
Each megasporophyll (carpel) is regarded as a modified leaf. It is about
12.7 cm to 25.4 cm long and can be divided into 3 parts: upper leafy portion,
middle ovule bearing portion and lower stalk. Ovules are formed on the lateral
side of the middle portion. The upper portion is pinnate and each pinna is
tapering to a point.
Two lateral rows of ovules are present on the lateral side of the middle
portion. In Cycas there is a great variation regarding the pinnate character of
megasporophyll and the number of ovules per sporophyll as a result of which in
various species of Cycas gradual reduction in megasporophylls can be traced.
The megasporophylls of C. revolula (Fig. 22A) are pinnate whereas those
of C. circinalis C. rumphii and C. beddomei (fig 22 B-D) are ovate lanceolate
structures. In C. pectinata and C. siamensis they are orbicular or rhomboidal
structures (Fig. 22B, F).
The laminar portion is well developed in C. revoluta, C.pectinata and C.
siamensis but reduced in C. circinalis, C. beddomei and C. rumphii (fig. 22).
The margin of lamina is serrate or dentate in C.circinalis, C.beddomei and C.
rumphii. The number of ovules differ in different species of Cycas. It is 1-6.
pairs in C.revoluta, C. Circinalis and only one pair in C. norambyana.
Megasporophylls are covered by yellow or brown hairs.
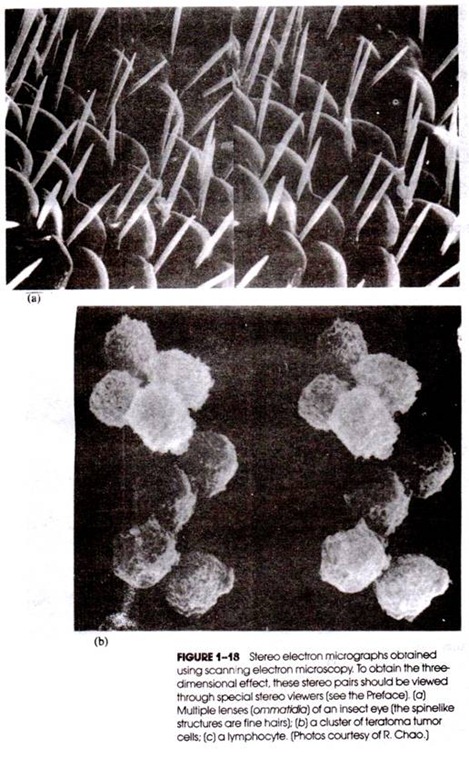
Structure of ovule (megasporangium):
The ovules are sessile and are borne laterally on the stalk. The ovules
of Cycas are largest in plant kingdom (7 cm long in C. thoursaii, 6 cm long x 4
cm diameter in C. circinalis) and can be seen by naked eye. The ovule is green
when young and is covered by hairs. At maturity its colour changes to orange
and hair also fall off.
The ovules are orthotropus (short and straight) and unitegmic (with one
integument).
The integument is very thick and
consists of three distinct layers:
(i) Outer, green or orange fleshy layer called outer sarcotesta
(ii) Middle, yellow stony layer called sclerotesta and
(iii) Inner fleshy layer or inner sarcotesta.
The parenchymatous tissue inside the integument is called nucellus. The
integument encloses all the nuclellus except at one point. This point or
opening is called micropyle. Just below the micropyle, the cells of the
nucellus form the nucellar beak.
Some of the cells of the nucellar beak dissolves and forms a cavity like
structure called pollen chamber. Just below the pollen chamber is present an
archegonial chamber. Micropyle leads into the pollen chamber. Just below the
floor of the archegonial chamber 3-6 archegonia are present towards the
micropylar end.
The ovule is supplied by three vasular traces (Fig. 23). The central
vascular trace enters the chalazal end of the nucellus. The inner and outer
vascular traces divide into two each, one branch supplies the outer fleshy
layer and the inner fleshy layer. Thus, the outer and inner fleshy layers
receive the vascular supply but the middle stony layers get no vascular supply
(Fig. 23).
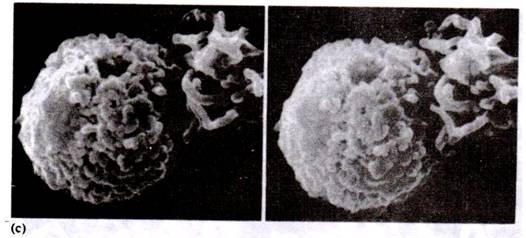
Development of Ovule:
When megasporophyll is young; in its middle portion 4-6 ovules arise as
a hypodermal mass of meristematic cells on the lateral side. These meristematic
cells divide and redivide to form a mass of parenchymatous cells known as
nucellus.
Soon the neighbouring cells at the base are also activated and they grew
upwards forming the integument which surrounds the nucellus on all sides except
at the top where a small opening is left which is known as micropyle (Fig. 24A,
B). In the beginning the nucellus and integument are free but afterwards due to
intercalary growth both of them fuse except in the region of micropyle.
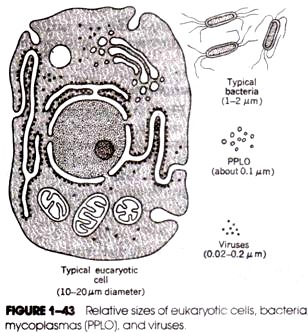
Deeply situated in the nucellus, any one cell enlarges and functions as
megaspore mother cell or embryo sac cell. It divides by meiosis to form a
linear tetrad of four megaspores. Upper three degenerate (Fig. 24 C-F) and
lower most is functional. It is the first cell of female gametophyte.
Gametophytic Phase:
Microspores are formed in microsporangia and megaspores are formed in
ovules after meiosis. Cycas is heterosporous because it produces two types of
spores.
Development of male gametophyte:
Each microspore is unicellular, unicelled structure with two layered
wall. Outer wall is known as exine while inner is known as intine.
The development of male gametophyte
takes place in two stages:
Stage I:
Development of male gametophyte before pollination: Development of male
gametopheyte or germination of pollen grains starts in situ i. e., they are
still inside the microsporangium.
First its nucleus divides into two, one of them goes towards the lower
side and is separated from the other by a cresent shaped wall resulting in the
formation of two unequal cells. The lower smaller cell is called as prothallial
cell and the upper bigger one as antheridial cell (Fig. 25 A, B).
The prothallial cell does not divide further but the antheridial cell
divides to form a generative cell (Fig. 25C) which is in close contact with the
prothallial cell and distal tube cell with a large nucleus. This stage is
called 3-celled stage consisting of a prothallial cell, generative cell and
tube cell.
At this stage microsporangia dehisce and the shedding of the pollen
grains takes place, (tapetum disintegrates, sporangium becomes dry, cells
shrink, sporangial wall ruptures radially at the line of dehiscence). Further
development of the pollen grains (II stage) takes place after pollination.

Development of Female Gametophyte:
The functional megospore (also called embryo sac cell or first cell of
female gametophyte) is haploid and it starts its development in situ i. e.,
within the nucellus (Fig 26 A). It absorbs the surrounding cells of the
nucellus and enlarges considerably. Its nucleus divides by free nuclear
divisions and as a result a large number of nuclei are formed. A vacuole
develops in the centre.
It pushes the free nuclei and cytoplasm of the megaspores towards its
periphery. (Fig. 26 B) Now the wall formation starts from periphery to the
centre (centripetal wall formation). It results in the formation of a cellular
tissue called endosperm or female prothallus or megagametophyte (Fig. 26 C).
The endosperm in Cycas is a haploid tissue formed before the fertilization.
Endosperm is nutritive in function. Simultaneously, a tiny space
develops on the upper side of the ovule between nucellus and the female
gametophyte due to degeneration of certain nucellar cells. This is called
archegonial chamber.
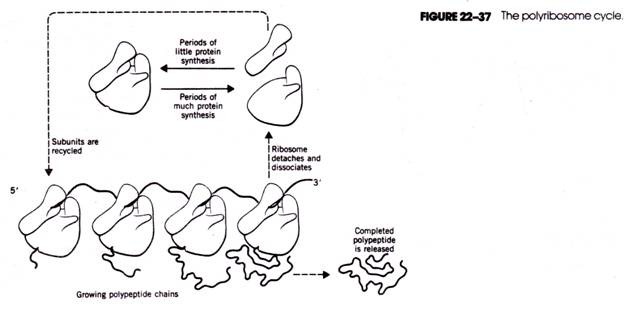
Development of archegonium:
The archegonia develop from the gemetophytic cells lining the
archegonial chamber towards the micropylar end. Any cell enlarges in size and
functions as archegonial initial (Fig. 27A). It divides transversely into an
upper primary neck cell and a lower central cell (Fig. 27 B). The primary neck
cell divides by a longitudinal division to form two neck cells. These cells
form the neck of the archegonium.
The central cell enlarges its size and its nucleus divides to form a
ventral canal nucleus and an egg nucleus (Fig.27 C). No wall is formed between
the venter canal nucleus and egg nucleus. Therefore, there is no neck canal
cell. Later on venter canal nucleus disorganises. The egg of the Cycas is
largest in all living plants measuring 5 mm in diameter.

Structure of Archegonium:
The mature archegonium consists of a neck of two neck cells. The
archegonial neck opens in the archegonial chamber. There is no neck canal cell.
There is no venter either. The egg and the venter canal nucleus remain
surrounded by the cells of the endosperm. These cells act as archegonium
jacket.
Pollination:
The pollination is anemophilous. The cells of nucellar beak present in
the pollen chamber disintegrate and form a viscous fluid. This fluid is
cohesive in nature. This fluid oozes out of the micropyle and collects in the
form of a pollination drop.
The pollen grains present in the air current at their 3-celled stage,
are entangled in the pollination drop. Gradually the pollen drop dries up and
the pollen grains are sucked into the pollen chamber through micropyle. Further
drying of this drop seals up the micropyle (Fig. 28) Pollen drop helps in collecting
the pollen grains at the micropyle in all gymnosperms.

II Stage. Development of male
gametophyte (after pollination):
After a definite period of rest (pollen grains may lie inside the pollen
chamber for quite some time say for four months, the interval between
pollination and fertilization is about 20 days in C. revoluta). Further
development of the male gametophyte takes place in the pollen chamber. The
exine breaks up and intine comes out in the form of pollen tube.
It may be branched or unbranched and acts as an absorbing organ (Pollen
tube acts only as an absorbing organ or hustorium because the sperms are formed
only after the complete development of the pollen tube) Pollen tube, penetrates
the nucellar tissue and comes to lie in the archegonial chamber. The generative
cell divides into a lower stalk cell and upper body cell (Fig. 25D). The stalk
cell does not divide further. The body cell divides at the time – micropyle of
fertilization.
Now two blepharoplasts appear, one at each pole of the nucleus of the
body cell in a transverse position. Body cell divides longitudinally into two
sperm cells, each of these having a single nucleus, blepharoplast and a small
amount of cytoplasm. Each sperm cell later on develops into a sperm and
blepharoplast gives rise to cilia. The sperms are liberated in the pollen tube
by the breaking of the sperm mother cell (Fig. 25E, F).
The sperms of the Cycas are largest in the plant kingdom (180-210 µ) and
visible to the naked eye. Each sperm is more or less triangular, top shaped
mobile structure having five to six spiral bands with thousands of cilia with a
single large nucleus. By means of their cilia, they move freely in the pollen
tube (Fig. 25F).
Fertilization:
At the time of fertilization, the nucellar tissue between the pollen
chamber and the archegonial chamber disorganise and simultaneously the venter
canal nucleus also disintigrates. The pollen tube reaches the archegonial
chamber (Fig. 29). The tip of the pollen tube ruptures releasing two male gametes
and fluid contents.
Due to this archegonial chamber becomes moist and the sperms move freely
in it with the help of cilia. Only a single sperm enters violently in each
archegonium through neck. Only the male nucleus of the sperm fuses with the egg
nucleus to form a zygote or oospore (2x). The fertilization in Cycas takes
place with the help of motile sperms.
This process is known as zooidogamy. It is accompanied by pollen tube
formation, a phenomenon known as siphonogamy. Sometimes more than one sperm enter
the archegonium but the male nucleus which first reaches near the nucleus
fertilizes the egg. Rest male nuclei degenerate. It is called polyspermy.

Embryogeny:
The fertilized egg, zygote or oospore is the first cell of the
sporophyte. The zygote contains dense cytoplasm and a large nucleus. It
enlarges in size and finally forms the embryo. In this whole process one year
time is utilised. The nucleus moves at the base and starts dividing by free
nuclear divisions to form about 256 free nuclei (Fig. 30A, B).
The nuclei and cytoplasm of the central region disorganise forming a
central vacuole (Fig. 30C). Subsequently wall formation takes place from the
base and advances towards the upper side forming a small mass of cells. This
embryonal mass of cells is called proembryo (Fig. 30D). It is meristematic in
nature. Some of the nuclei in the upper region remain without cell walls.
The proembryo soon gets differentiated
into three zones (Fig. 30E):
(1) Upper haustorial zone at the micropylar end
(2) The middle suspensor zone
(3) Tha basal embryonal zone.
The haustorial zone absorbs the nutrition for the developing embryo.
Suspensor zone elongates considerably forming a long, spirally coiled
suspensor, which pushes the embryo in the food containing cells of the
endosperm. The embryonal zone is differentiated into embryo. It is
differentiated into two cotyledons (three in C. circinalis).
A plumule is then differentiated in between the cotyledonary depression.
The embryonal part lying below the cotyledonary attachment is the hycotyl. A
radicle is now differentiated at the base of the hypocotyl. It is protected by
a hard pad like protective covering called coleorhiza. It provides protection
to the radicle (Fig. 30 F, G).
Several archegonia may be fertilized in an ovule of Cycas. It results in
the formation of several zygotes. These zygotes may undergo initial development
but usually one embryo reaches maturity.

Structure of seed:
After fertilization, the ovule is transformed into seed. The nucellus
and the inner layer of integument are used up as nourishment by developing
embryo. The mature seed appears as orange-red or reddish brown structure.
It comprises the following structures:
a. Testa or seed coat:
It is formed by the outer brightly coloured fleshy layer and the middle
layer of the integuments.
b. Micropyle:
It is present in the form of small opening at the top of the seed.
c. Endosperm:
Inner to the seed coat lie the wall tissues called endosperm. The cells
store a large amount of food material.
d. Embryo:
Embedded in the endosperm lies the embryo. It consists of two
cotyledons, plumule and radicle. The embryo remains suspended in the endosperm
by a long spirally coiled suspensor (Fig. 32).

Thus, a mature seed of Cycas represents
three generations:
Seed Coat:
It is formed by the integument and represents parent-sporophytic
generation.
Endosperm:
Represents the gametophytic generation.
Embryo:
It represents the new sporophytic generation.
Testa is sweet in taste and emits pleasant odour. The two
characteristics i. e., red colour and pleasant odour are responsible for their
zoochorus (orinthochorous) dispersal.
Germination of seed:
Seeds remain viable for not more than a few months. Under suitable
conditions the seed starts germination. It absorbs water and embryo expands.
The expansion of the embryo breaks open the hard seed coat.
The coleorhiza protrudes out and is pierced by the growing radicle which
grows down and forms the tap root or primary root (Fig. 32A). The cotyledons do
not come out of the seed coat but they absorb food from the endosperm for the
growing embryo.
The stalk of the cotyledons elongates to carry the plumule out of the
seed coat. The growing plumule at first, forms a few scaly leaves and then
foliage leaves. The young foliage leaves show circinate vernation (Fig. 32
B-D).
The seed germination is hypogeal because the cotyledons remain
underground enclosed in the female prothallus or endosperm. However, according
to some workers cotyledons do not come out of the seed during germination as
they are haustorial in nature, but their bases remain exposed, hence the seed
germination may be said to epigeal.
Economic Importance of Cycas:
1. Several species of Cycas of e.g., C. revoluta, C. circinalis C.
bedomei are grown in the gardens as ornamental plants.
2. From the stem ‘sago’ a kind of starch, is obtained. Hence, it is also
called sago palm (C. revoluta)
3. Leaves of C. circinalis are used to prepare hats, baskets and mats.
The leaves of Cycas are extensively used for decorative purposes and floral
decoration.
4. The resin obtained from C. rumphii is applied to malignant ulcers.
5. Juice of tender leaves of Cycas is useful for vomiting and flatulence
(the presence of excessive gas in the digestive tract).
6. Pollens of C. circivalis are narcotic. Its seeds ground to paste with
coconut oil are useful for sores and swellings.
7. Seeds of Cycas are roasted and are used as food in Assam and certain
islands.
8. The seeds and stem of C. revoluta are used in making wire in Japan.
9. The flour of seeds of Cycas is called Indum Podi and is used in the
preparation of cakes and porridges.
10. The young succulent leaves of Cycas are also cooked as vegetable.


{kind=link}
0 Comments